admin
Admin

Nombre de messages : 2238
Date d'inscription : 27/10/2008
 |  Sujet: Vers un nouveau traitement du cancer et des maladies génétiques Sujet: Vers un nouveau traitement du cancer et des maladies génétiques 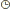 Jeu 8 Oct - 21:37 Jeu 8 Oct - 21:37 | |
| Actuellement la plupart des maladies génétiques sont incurables. En 2000 puis plus récemment en juin 2002, la thérapie génétique a permis de soigner des "bébés bulles". Cependant ces protocoles ont récemment été arrétés suite à l'apparition chez un des enfants traités d'une leucémie, liée à l'intégration du transgène dans un gène suppresseur de tumeur. Ce problème souligne la difficulté de mettre au point une technique efficace et sure de thérapie génique. Ce dossier aborde la possibilité d'utiliser une autre approche thérapeutique afin de soigner ou de réduire les effets d'un nombre important de maladies génétiques et de cancers.  Image de la sous unité 50S du ribosome de Haloarcula marismortui à une résolution de 2,4Å. L'ARNr est représenté en gris, les protéines constitutives du ribosome sont représentées en jaune et se situent quasiment uniquement à la périphérie. D'après Ban, N et collaborateurs. Science, 289, 905-920 (2000)1.Les mécanismes d'expression des gènesDeux grandes étapes sont nécessaires afin d'obtenir une protéine à partir d'un gène. La transcription permet d'obtenir une copie du gène présent sur le chromosome, sous forme d'un ARN messager (ARNm). La traduction est l'étape durant laquelle cet ARNm est lu par le ribosome Image de la sous unité 50S du ribosome de Haloarcula marismortui à une résolution de 2,4Å. L'ARNr est représenté en gris, les protéines constitutives du ribosome sont représentées en jaune et se situent quasiment uniquement à la périphérie. D'après Ban, N et collaborateurs. Science, 289, 905-920 (2000)1.Les mécanismes d'expression des gènesDeux grandes étapes sont nécessaires afin d'obtenir une protéine à partir d'un gène. La transcription permet d'obtenir une copie du gène présent sur le chromosome, sous forme d'un ARN messager (ARNm). La traduction est l'étape durant laquelle cet ARNm est lu par le ribosome, pour obtenir une protéine. Afin d'obtenir une protéine active chacune de ces deux étapes doit être contrôlée rigoureusement pour être effectuée fidèlement.* La traduction Longtemps négligée dans l'étude de la régulation des gènes, au profit de la transcription, la traduction a fait l'objet ces dernières années d'études intensives. La traduction correspond à la lecture de l'ARNm par le ribosome. Cette lecture s'effectue trois nucléotides par trois nucléotides (3 nucléotides = 1 codon). Elle commence au codon initiateur (AUG), pour se terminer à l'un des trois codons stop (UAA, UAG ou UGA) (Figure 1). Au niveau de chaque codon, le ribosome incorpore un ARN de transfert (ARNt) chargé à son extrémité avec un acide aminé. Cet ARNt possède une séquence complémentaire (anticodon) au codon présent dans le site de décodage du ribosome. L'appariement ARNt-ARNm permet de distinguer l'ARNt qui doit effectivement être incorporé d'un ARNt entré dans le ribosome par erreur (Figure 2). Une fois l'identité de l'ARNt vérifiée, la chaîne protéique en élongation est transférée sur le nouvel acide aminé, grâce à l'activité peptidyl-transferase du ribosome. Une fois le transfert effectué, le ribosome se déplace d'un codon (translocation) et un nouveau cycle s'effectue jusqu'à ce que le ribosome rencontre un codon stop (figure 1). Lorsque le codon stop pénètre dans le ribosome, les facteurs de terminaison (eRF) vont provoquer l'arrêt de la synthèse protéique, le relargage de la protéine nouvellement synthétisée, et la dissociation des deux sous-unités du ribosome (figure 1).  Schéma général représentant les 5 grandes étapes de la traduction Schéma général représentant les 5 grandes étapes de la traduction Représentation schématique des sites A et P du ribosome. L'ARNm est repésenté en bleu, les ARNt en noir, les acides aminés par des sphères colorées. L'ARNt pénètre dans le ribosome au site A. Si l'appariement codon/anticodon est correct, alors il y a transfert de la protéine en élongation sur le nouvel acide aminé apporté par l'ARNt. Dans le cas contraire, l'ARNt est éjecté du ribosome et un autre prend sa place.La traduction est un point de régulation important de l'expression des gènes (1). Il a été montré que ces régulations pouvaient se dérouler au niveau de l'initiation de la traduction, de l'élongation (2, 3), mais aussi au niveau de la terminaison et ceci aussi bien chez les eucaryotes que les procaryotes (4, 5). * La terminaison de la traduction. Les mécanismes moléculaires de cette dernière étape de l'expression des gènes ont récemment été élucidés chez les bactéries (6). En revanche chez les eucaryotes ces mécanismes demeurent encore mal compris. Dans les conditions normales l'efficacité de terminaison est de l'ordre de 99,7 %. Cependant il a clairement été démontré que cette efficacité était fortement dépendante de plusieurs paramètres Représentation schématique des sites A et P du ribosome. L'ARNm est repésenté en bleu, les ARNt en noir, les acides aminés par des sphères colorées. L'ARNt pénètre dans le ribosome au site A. Si l'appariement codon/anticodon est correct, alors il y a transfert de la protéine en élongation sur le nouvel acide aminé apporté par l'ARNt. Dans le cas contraire, l'ARNt est éjecté du ribosome et un autre prend sa place.La traduction est un point de régulation important de l'expression des gènes (1). Il a été montré que ces régulations pouvaient se dérouler au niveau de l'initiation de la traduction, de l'élongation (2, 3), mais aussi au niveau de la terminaison et ceci aussi bien chez les eucaryotes que les procaryotes (4, 5). * La terminaison de la traduction. Les mécanismes moléculaires de cette dernière étape de l'expression des gènes ont récemment été élucidés chez les bactéries (6). En revanche chez les eucaryotes ces mécanismes demeurent encore mal compris. Dans les conditions normales l'efficacité de terminaison est de l'ordre de 99,7 %. Cependant il a clairement été démontré que cette efficacité était fortement dépendante de plusieurs paramètres : A. La nature du codon stop. Les trois codons stop ne permettent pas la même efficacité de terminaison. Il est généralement admis que le codon UAA est le plus efficace, les codons UGA et UAG présentant une efficacité de terminaison légèrement plus faible. Il n'existe pas naturellement dans la cellule des ARNt possédant un anticodon parfaitement adapté pour reconnaître les codons stop. Cependant, l'hypothèse la plus probable pour expliquer ces différences d'efficacité entre les différents codons stop est l'existence dans la cellule d'un nombre plus ou moins important d'ARNt naturels pouvant être incorporés au niveau du codon stop par le ribosome (même si la reconnaissance ne s'effectuera pas par un appariement complet codon/anticodon). B. Rôle du contexte nucléotidique du codon stop. Une analyse statistique du contexte nucléotidique des codons stop présents à la fin des gènes chez divers organismes a montré un biais significatif des nucléotides situés en amont et en aval du codon stop. L'influence des deux codons situés en amont du codon stop a lieu principalement au travers des acides aminés incorporés. Ces acides aminés interagissent avec le tunnel de sortie du ribosome et modifient probablement ainsi l'affinité du ribosome pour les facteurs de terminaison.
Dans tous les organismes étudiés le nucléotide situé immédiatement après le codon stop présente un très fort biais de représentation. Ce résultat a entraîné certains scientifiques à émettre l'hypothèse que le signal d'arrêt de la traduction ne serait pas un triplet mais un quadruplet (7). Cependant, même si de nombreuses expériences ont démontré l'importance de ce 4ème nucléotide pour l'efficacité de terminaison de la traduction, aucune preuve n'a jamais été apportée quant à un signal composé de 4 nucléotides au lieu de trois. De plus, aucune règle générale n'a pu être déduite des différentes études réalisées car la nature du nucléotide dépend du codon stop ainsi que de l'organisme étudié. D'autres travaux ont mis en évidence que le contexte influençant l'efficacité de terminaison de la traduction était encore plus étendu (6-9 nucléotides) que ce qui avait été préalablement estimé (8, 9). Plusieurs hypothèses ont été avancées pour expliquer le rôle du contexte nucléotidique situé en aval du codon stop (appariement avec l'ARN ribosomique, interaction avec des protéines du ribosome), mais aucune preuve expérimentale n'a encore été apportée pour confirmer ou infirmer ces hypothèses. Que se passe-t-il lorsque le ribosome ne s'arrête pas au codon stop?Nous venons de voir que la présence d'un codon stop ne signifie pas systématiquement l'arrêt de la synthèse protéique. En fonction de la nature du codon stop et du contexte nucléotidique il est possible d'atteindre plus de 30% de passage du codon stop (suppression, ou translecture) chez la levure Saccharomyces cerevisiae. Après avoir franchi le codon stop, le ribosome continue la traduction (dans la même phase) jusqu'au codon stop suivant. Cela aboutit à la synthèse d'une protéine possédant une extension carboxy-terminale. Cette extension peut avoir plusieurs conséquences : modifier la stabilité de la protéine, modifier son activité, ou la localisation de la protéine dans la cellule. Il est aussi possible que cette extension possède une nouvelle activité enzymatique. Ce dernier cas est fréquemment retrouvé dans l'expression de gènes viraux qui utilisent un mécanisme de translecture pour exprimer deux protéines à partir du même ARNm. C. Les drogues. Le ribosome est une cible privilégiée pour de nombreuses drogues notamment les antibiotiques. Ceux-ci, en inhibant la synthèse protéique des bactéries, permettent de lutter efficacement contre les infections. Le ribosome eucaryote est structuralement bien différent du ribosome procaryote. Ceci implique donc que les molécules actives sur le ribosome procaryote n'auront pas le même effet (voir pas d'effet du tout) sur le ribosome eucaryote. Certains antibiotiques ont la particularité de modifier la fidélité de la traduction, en rendant le ribosome moins fidèle. Nous allons maintenant présenter dans le chapitre suivant une classe particulière d'antibiotiques : les aminoglycosides.2.Une classe d'antibiotiques particulière : les aminoglycosidesLes aminoglycosides sont utilisés de manière courante à forte concentration pour traiter les infections bactériennes. Il a été développé ces dernières années un grand nombre de molécules naturelles ou synthétiques de cette famille. Cependant notre intérêt pour les aminoglycosides ne réside pas dans leur capacité à traiter les infections bactériennes, mais dans leur capacité à augmenter les erreurs de lecture du ribosome au cours de l'élongation et de la terminaison. Les antibiotiques les mieux décrits pour induire des erreurs lors de la terminaison de la traduction sont la gentamicine, le G418 et la paromomycine (10, 11). Plus récemment il a été mis en évidence un effet similaire avec deux autres antibiotiques: la tobramycine et l'amikacine (12). Cependant quelques membres de cette famille n'ont aucun effet mesurable sur l'efficacité de terminaison de la traduction (Kanamycine, HygromycineB). Les raisons pour lesquelles certains antibiotiques sont efficaces et d'autres pas demeurent encore incertaines, d'autant que les différences structurales sont parfois assez faibles (voir structure des antibiotiques). * Structure des aminoglycosides Il existe deux classes principales d'aminoglycosides: ceux qui possèdent un groupe déoxystreptamine 4,6 disubstitué (Figure 3A) et ceux possédant un groupe déoxystreptamine 4,5 disubstitué (Figure 3B). Le G418, la sisomycine, ainsi que la gentamicine font partie de la famille 4,6 disubstituée. Il faut noter que la gentamicine commercialement disponible est un mélange de trois formes (C1a, C1 et C2) différentes les unes des autres par le groupement attaché au carbone 6 du cycle I. Quatre autres membres de cette famille, Kanamycine A, B, tobramycine, et amikacine, possèdent en plus un cycle III différents. Les membres de la famille 4,5 disubstituée diffèrent de la famille précédente par deux caractéristiques principales. Tout d'abord ils contiennent une liaison speciale entre le cycle II et le cycle III, ensuite ils sont fréquemment constitués de 4 cycles ou plus. La paromomycine fait partie de cette deuxième famille d'aminoglycosides.  Représentation des différentes familles d'aminoglycosides. A: famille 4,6 disubsituée. B: famille 4,5 disubsituée. C: d'autres aminoglycosides* Mode d'action et spécificité de ces antibiotiques. Ces antibiotiques interagissent avec le site de décodage du ribosome et induisent un changement de conformation du site de décodage. Ce changement de conformation diminue la capacité du ribosome à distinguer un ARNt correct (appariement codon-anticodon parfait) d'un ARNt incorporé par erreur. Ainsi en présence de ces antibiotiques le ribosome va incorporer plus efficacement le mauvais ARNt. Cette diminution de la fidélité du ribosome va permettre une meilleure incorporation des ARNt au niveau des codons stop, et ainsi augmenter l'efficacité de translecture. Cependant il a été démontré indépendamment par deux équipes Américaines que l'efficacité de suppression de ces antibiotiques était fortement dépendante du codon stop et de son contexte nucléotidique (principalement le nucléotide suivant immédiatement le codon stop) (13, 14). Il est actuellement difficile d'expliquer pourquoi l'effet de ces antibiotiques est tellement dépendant du codon stop et de son contexte. Il est donc important de noter, que lorsqu'une cellule est traitée par un de ces antibiotiques, tous les codons stop ne vont pas être supprimés avec la même efficacité. Ceci permet à la cellule de continuer d'exprimer la plupart des ses gènes de manière tout à fait normale. Représentation des différentes familles d'aminoglycosides. A: famille 4,6 disubsituée. B: famille 4,5 disubsituée. C: d'autres aminoglycosides* Mode d'action et spécificité de ces antibiotiques. Ces antibiotiques interagissent avec le site de décodage du ribosome et induisent un changement de conformation du site de décodage. Ce changement de conformation diminue la capacité du ribosome à distinguer un ARNt correct (appariement codon-anticodon parfait) d'un ARNt incorporé par erreur. Ainsi en présence de ces antibiotiques le ribosome va incorporer plus efficacement le mauvais ARNt. Cette diminution de la fidélité du ribosome va permettre une meilleure incorporation des ARNt au niveau des codons stop, et ainsi augmenter l'efficacité de translecture. Cependant il a été démontré indépendamment par deux équipes Américaines que l'efficacité de suppression de ces antibiotiques était fortement dépendante du codon stop et de son contexte nucléotidique (principalement le nucléotide suivant immédiatement le codon stop) (13, 14). Il est actuellement difficile d'expliquer pourquoi l'effet de ces antibiotiques est tellement dépendant du codon stop et de son contexte. Il est donc important de noter, que lorsqu'une cellule est traitée par un de ces antibiotiques, tous les codons stop ne vont pas être supprimés avec la même efficacité. Ceci permet à la cellule de continuer d'exprimer la plupart des ses gènes de manière tout à fait normale.
Dans les conditions normales d'expression des gènes le ribosome rencontre le codon stop à la fin de la séquence codante. Seulement ce n'est malheureusement pas toujours le cas et il arrive que la synthèse protéique s'arrête prématurément en raison de l'apparition d'un codon stop.3.Qu'est-ce qu'une maladie génétique Les maladies génétiques sont toutes dues à une modification de la séquence initiale d'un gène (ou mutation). Il existe plusieurs catégories de mutation: -Les mutations "larges" qui peuvent toucher de très grandes régions des chromosomes (délétions, insertions, inversions),- des mutations "ponctuelles" qui ne modifient qu'un seul nucléotide. Ces dernières peuvent modifier la composition en acide aminé d'une protéine (mutations faux-sens), ou interrompre prématurément la synthèse protéique (mutations non-sens) en faisant apparaître un codon stop au milieu d'un gène. L'apparition d'un codon stop dans un gène est souvent un événement délétère pour la cellule. Ainsi 18% des mutations touchant le gène DYS et causant la dystrophie musculaire de Duchenne, aussi appelée myopathie de Duchenne (maladie neuromusculaire touchant 1 garçon sur 3500), sont des mutations non-sens. Une proportion importante des cancers est aussi due à l'apparition d'une mutation non-sens, ainsi 10 % des mutations inactivant le gène de la p53 (gène suppresseur de tumeur) sont des mutations non-sens, et ce gène est retrouvé muté dans plus de 50 % des cancers humains. De même, près de 30 % des mutations inactivant le gène APC (gène suppresseur de tumeur du colon, retrouvé muté dans 80% des cancers du colon), sont des mutations non-sens. Dix huit pour cent des mutations identifiées dans le gènes BRCA1 et 12% dans le gène BRCA2 sont des mutations non-sens (15). Or ces deux gènes sont retrouvés mutés dans une grande majorité des cancers héréditaires du sein. En plus des différents cancers, ce sont plus de 200 maladies génétique différentes qui sont attribuées à l'apparition d'un codon non-sens.
A la lecture de ces chiffres on constate immédiatement que, s'il était possible de reprogrammer le ribosome afin qu'il ne s'arrête pas aux codons stop apparus prématurément dans le gène, alors on pourrait soigner un grand nombre de maladies actuellement incurables en rétablissant la synthèse d'une protéine active.4.Principe thérapeutiqueIl s'agit donc d'utiliser les propriétés de certains aminoglycosides afin d'éviter que le ribosome ne s'arrête au codon stop apparu par mutation. Ainsi, il devrait être possible de rétablir l'expression de la protéine, et ce malgré la présence de la mutation non-sens. Si le principe de base de cette nouvelle stratégie thérapeutique paraît simple, celle-ci n'en demeure pas moins compliquée à mettre en œuvre comme nous allons le voir dans les paragraphes suivants5.Les résultats déjà obtenusRésultats in vitro et ex-vivo De nombreuses études ont été réalisées in vitro et ont démontré la validité de l'approche. Ainsi il a été montré que les aminoglycosides: gentamicine, tobramycine, amikacine et G418 étaient capables de restaurer l'expression de gènes inactivés par un codon stop. Cependant l'efficacité de ces antibiotiques est extrêmement dépendante du contexte nucléotidique entourant le codon stop. De plus les résultats publiés sont parfois contradictoires, ce qui rend difficile l'interprétation du rôle du contexte nucléotidique dans l'efficacité de suppression des codons stop.
Des résultats similaires ont été obtenus ex-vivo (dans des lignées de cellules en culture), sur des mutations touchant les gènes p53, CFTR, IDUA, DYS.
La plupart des systèmes utilisés pour quantifier l'effet de ces antibiotiques sont des systèmes reporters dans lesquels a été clonée une séquence plus ou moins grande encadrant la mutation non sens. Cette approche bien que très précieuse (et quasi irremplaçable) pour déterminer l'efficacité d'un antibiotique à supprimer un codon stop se heurte à un problème majeur. Elle ne permet pas de déterminer si la protéine exprimée par la suppression du codon stop sera active ou non. Nous avons vu que l'antibiotique favorisait l'incorporation d'un ARNt au niveau du codon stop. Cet ARNt va donc permettre de continuer la traduction jusqu'au codon stop suivant et restaurer ainsi l'expression de la protéine. Mais ensuite celle-ci doit être active! Or il est tout à fait possible que l'acide aminé incorporé via l'ARNt au niveau du codon stop modifie fortement la structure, l'activité ou la stabilité de la protéine enpêchant ainsi son fonctionnement normal.
Cependant certains chercheurs ont tiré profit de l'activité enzymatique de la protéine affectée par la mutation stop pour étudier directement l'effet des antibiotiques sur l'activité de la protéine elle même. La protéine étudiée (TPPI) est codée par le gène CLN2. C'est une protéase lysosomiale (tripeptidyl-peptidase) qui, lorsqu'elle est inactive provoque une maladie neurodégénérative fatale des enfants atteints (espérance de vie de 8 à 15 ans) (16). Ce gène est particulièrement bien adapté à l'étude des aminoglycosides pour trois raisons. 1. La protéine possède une activité enzymatique quantifiable.
2. Certains cas cliniques ont montré que des patients possédant entre moins de 1% et 5% d'activité enzymatique ont une durée de vie rallongée (20 à 40 ans), que la maladie apparaissait plus tardivement (6-8 ans au lieu de 2-4 ans) et que souvent les symptômes étaient moins sévères.
3. Dans plus de 50% des cas, le gène est inactivé par la présence d'une mutation non-sens et comme actuellement aucun traitement n'existe contre cette maladie, la mise au point d'une thérapie à base d'aminoglycosides pourrait améliorer significativement la vie de certains malades Coupe en immunohistochimie de myotudes (cellules musculaires) provenant de souris saines ou de souris mdx. La présence de dystrophine est révélée par un anticorps dirigé contre la partie terminale de la protéine. Le marqueur fluorescent rouge utilisé pour cette détection est la rhodamine. On constate très clairement la réapparition de la dystrophine dans les cellules traitées à la gentamicineLes études ont portées sur trois mutations différentes. Les résultats ont montré aucune suppression de la mutation Gln66. En revanche l'addition de 60 microgramme/ml de gentamicine a permis de retrouver 8% d'activité TPPI dans des cellules dont le gènes CLN2 possédait les mutations Arg127 et Arg208 (cellules hétérozygotes), enfin 0.5% d'activité ont été mesuré après traitement à la gentamicine dans les cellules possédant seulement la mutation Arg208 (17). Les auteurs expliquent que ces différences peuvent provenir de différents paramètres, tels que la stabilité de l'ARNm (les ARNm contenant un codon stop précoce sont instables), la durée de vie de la protéine etc…Quoiqu'il en soit ces résultats montrent qu'il est possible de restaurer l'activité d'une protéine par l'utilisation de la gentamicine. Cette restauration n'est pas complète, mais est significative surtout si on prend en compte qu'il peut suffire de quelques % d'activité pour annuler l'effet de la mutation stop. Coupe en immunohistochimie de myotudes (cellules musculaires) provenant de souris saines ou de souris mdx. La présence de dystrophine est révélée par un anticorps dirigé contre la partie terminale de la protéine. Le marqueur fluorescent rouge utilisé pour cette détection est la rhodamine. On constate très clairement la réapparition de la dystrophine dans les cellules traitées à la gentamicineLes études ont portées sur trois mutations différentes. Les résultats ont montré aucune suppression de la mutation Gln66. En revanche l'addition de 60 microgramme/ml de gentamicine a permis de retrouver 8% d'activité TPPI dans des cellules dont le gènes CLN2 possédait les mutations Arg127 et Arg208 (cellules hétérozygotes), enfin 0.5% d'activité ont été mesuré après traitement à la gentamicine dans les cellules possédant seulement la mutation Arg208 (17). Les auteurs expliquent que ces différences peuvent provenir de différents paramètres, tels que la stabilité de l'ARNm (les ARNm contenant un codon stop précoce sont instables), la durée de vie de la protéine etc…Quoiqu'il en soit ces résultats montrent qu'il est possible de restaurer l'activité d'une protéine par l'utilisation de la gentamicine. Cette restauration n'est pas complète, mais est significative surtout si on prend en compte qu'il peut suffire de quelques % d'activité pour annuler l'effet de la mutation stop.
Des résultats publiés en janvier 2002 par une équipe américaine montrent qu'il est possible de supprimer des mutations du gène p53 dans des cellules fibroblastiques embryonnaires de rats (13). Grâce à une quantification précise de l'efficacité de suppression, mais aussi grâce à l'utilisation d'un test indirect de l'activité de la protéine p53, ils ont montré que les cellules traitées avec 200g/ml de gentamicine, ou 800mg/ml d'amikacine possédaient une activité p53 significative. Ces résultats sont très importants car ils démontrent que la protéine p53 qui est une cible majeure de mutations entraînant des cancers chez l'homme peut récupérer une partie de son activité après un traitement à la gentamicine ou l'amikacine.Résultats in-vivo Les résultats les plus prometteurs à ce jour, ont été obtenus chez la souris. Il existe une mutation chez la souris (mutation mdx) qui sert de modèle animal à la myopathie de Duchenne. Le gène affecté par la mutation est le gène codant la dystrophine musculaire, qui est une protéine de structure des muscles. Les signes de la maladie apparaissent vers 3 ans, petit à petit une faiblesse musculaire apparaît. Cette faiblesse musculaire atteint les muscles moteurs, mais aussi les muscles respiratoires et le cœur. La membrane des cellules musculaire étant fragilisée, elle ne résiste pas aux efforts de contractions. Les cellules se détruisent relargant dans le sang une enzyme musculaire la Créatine Kinase (CK) qui est mesurable.
Les souris mdx possèdent une mutation non-sens dans le gène codant la dystrophine résultant d'une absence de dystrophine dans les muscles. Les chercheurs ont traité les souris avec de la gentamicine, soit par injection, soit en continu par un mécanisme de pompe. Les résultats montrent qu'après le traitement à la gentamicine, il est de nouveau possible de mettre en évidence la présence de la dystrophine dans les muscles (10 à 20% de la quantité normale), que ceux-ci sont moins fragiles, qu'une certaine activité est retrouvée et que le niveau de CK dans le sang est significativement diminué indiquant une meilleure résistance des cellules musculaires (18). Cependant les auteurs notent que la concentration en gentamicine diminue très rapidement pour atteindre 4 heures après l'injection une concentration très faible (1g/ml) qui est maintenue pendant 24h. Cette rapidité de métabolisation de l'antibiotique peut être un problème pour son efficacité.
Si ces résultats sont très encourageants il convient de les relativiser. Tout d'abord, d'autres équipes ont échoué en tentant de reproduire ces résultats. Ceci peut être expliqué par l'utilisation de lots de gentamicine ayant des compositions différentes (la gentamicine disponible commercialement est un mélange de trois formes de gentamicine, qui varient en proportion). Or comme aucune solution de gentamicine pure n'existe actuellement, il n'est pas possible de déterminer lequel des ces trois composants est efficace. Ensuite, la mutation stop présente chez la souris mdx est UAAA, qui d'après d'autres chercheurs ne permet pas plus de 1% de suppression du codon stop dans les cellules en culture. Or les auteurs reportent 10 à 20% de dystrophine chez la souris. Il n'est donc pas impossible que la restauration de l'expression de la protéine se fasse par un autre mécanisme que la suppression du codon stop chez la souris (un épissage alternatif de l'ARN pré-messager par exemple).
Actuellement trois essais cliniques chez l'homme ont été entrepris afin de déterminer si la gentamicine pouvait être utilisée. Ces trois essais ont donnés des résultats opposés. Les deux premiers ont montré que l'administration de gentamicine chez des patients possédant une mutation non-sens dans le gène CFTR permettait de retrouver partiellement l'expression de la protéine ainsi qu'une activité dans l'éptihélium nasal (11, 19). En revanche le troisième essai n'a montré aucune amélioration de patients atteints de la myopathie de Duchenne et traités à la gentamicine (20). Dans cet essai, il n'a pas été non plus possible de mettre en évidence la présence de la forme complète de la dystrophine.
Il est important de noter qu'il existe une différence essentielle, qui pourrait permettre d'expliquer ces différences de résultats. La protéine CFTR possède une activité enzymatique, alors que la dystrophine est une protéine de structure des muscles. Il est facilement envisageable que très peu d'une protéine ayant une activité enzymatique permette d'attenuer les effets d'une maladie, alors qu'il faudra une quantité plus importante d'une protéine de structure. Si cette hypothèse est valable, alors les maladies provenant de la disparition d'une activité enzymatique (CFTR, cancer(p53), IUDA …) seront de meilleurs cibles thérapeutiques.6.Espoirs et limites de cette approche thérapeutiqueComme nous venons de le voir, la possibilité de soigner certaines maladies génétiques en administrant des aminoglycosides peut être source de grands espoirs, tellement cette solution thérapeutique peut paraître simple. Seulement elle se heurte encore à de nombreux problèmes. Le premier est le faible taux de suppression observé chez l'animal et les cellules en culture. Le second est que l'effet des antibiotiques est très variable d'une mutation à l'autre. Ensuite il n'est pas envisageable de traiter les malades à vie avec les antibiotiques, en raison des dérèglements que cela engendrerait pour la flore intestinale ainsi que pour les reins chargés d'éliminer l'antibiotique. De plus il a été rapporté que que la gentamicine avait parfois comme effet secondaire une surdité plus ou moins importante. Nous avons aussi vu, qu'actuellement il semble que les antibiotiques utilisés soient très rapidement métabolisés par l'organisme. Pour être réellement efficace, et sans contraindre le patient à être sous perfusion 24H/24, il faudrait des molécules plus stables.
Enfin, l'un des obstacles les plus sérieux à l'utilisation des antibiotiques pour supprimer les mutations stop, est qu'il est essentiel d'assurer un bon dosage. Il est effectivement important d'avoir une concentration d'antibiotique suffisante pour permettre le maximum de suppression de la mutation stop. Mais à trop forte concentration l'antibiotique risque d'avoir l'effet inverse, en effet nous avons vu que ces antibiotiques provoquaient aussi des erreurs lors de l'élongation (il y a donc incorporation d'un mauvais acide-aminé dans la protéine). Si ce phénomène (appelé misincorporation) est lui aussi trop important, nous allons provoquer la synthèse de nombreuses protéines anormales, sans pouvoir prédire quels seront leurs effets sur la cellule. Il est possible que ce problème ait été partiellement résolu récemment par une équipe américaine. Il viens d'être publié au mois de septembre 2002, qu'une nouvelle classe d'antibiotique, les oxazolidinones, qui se lient à une des sous-unités du ribosome, était capable d'induire la suppression des codons stop sans pour autant augmenter le nombre d'erreur du ribosome lors de l'élongation (21). Ces résultats n'ont pour l'instant été obtenus que dans les bactéries, il reste donc à les confirmer chez les mammifères.
Les futures recherches devront s'orienter vers la recherche de molécules plus efficaces dans la suppression des codons stop, tout en ayant des effets secondaires moins importants (probablement en n'affectant pas le taux d'erreur du ribosome pendant l'élongation). Cette voie de recherche est très prometteuse car elle devrait permettre une amélioration sensible de la qualité de vie et de l'espérance de vie des malades actuellement incurable | |
|